Alteration of hepatic anti-oxidant systems by 4-nonylphenol, a metabolite of alkylphenol polyethoxylate detergents, in Far Eastern catfish Silurus asotus
Article information
Abstract
Objectives
This study aimed to estimate the effects of 4-nonylphenol (NP), a ubiquitously present surfactant in aquatic environments, on the anti-oxidant systems of the liver in the Far Eastern catfish Silurus asotus.
Methods
Changes in biochemical parameters involved in glutathione (GSH)-related and other anti-oxidant systems were analyzed following 4 weeks of 4-NP administration (0.1 and 1.0 mg/kg diet) via a formulated diet to catfish.
Results
4-NP exposure induced an elevation in hepatic lipid peroxide levels and an accompanying decrease in reduced state GSH after 2 weeks, suggesting pro-oxidant effects of the chemical in catfish. This oxidative stress was associated with an inhibition of the GSH-utilizing enzyme glutathione peroxidase at the same time point. This inhibition was restored after 4 weeks. The activities of other anti-oxidant enzymes, i.e., glutathione reductase, superoxide dismutase and catalase were increased after 4 weeks. These enzyme increases occurred more strongly at the higher 4-NP concentration (1.0 mg/kg diet).
Conclusions
4-NP given to catfish at 0.1 to 1.0 mg/kg diet, concentrations relevant to environmental levels, depletes the endogenous anti-oxidant molecule GSH and temporarily inhibits GSH-related anti-oxidant enzymes. Such declines in anti-oxidant capacity and elevated oxidative stress seem to be compensated eventually by subsequent activation of various anti-oxidant enzyme systems.
Introduction
4-Nonylphenol (4-NP) is a typical endocrine disruptor which originates as a result of the microbial metabolism of nonylphenol polyethoxylates that are widely used as surfactants [1,2]. The large volume of production and use of surfactants has led to widespread release into the aquatic environment [3]. Although 4-NP is the degradation product of surfactants, this metabolite is more toxic than its parent compound and degradation proceeds very slowly in aquatic environments. The physicochemical characteristics of 4-NP can also lead to bio-concentration and bio-accumulation in aquatic organisms [4].
Clearly, the best documented 4-NP toxicity in aquatic organisms is the perturbation of endocrine functions [5]. Due to its chemical similarity to estrogens, this contaminant binds to estrogen receptors, leading to vitellogenin expression in the fish liver [6,7].
It has been recently demonstrated that 4-NP stimulates the production of reactive oxygen species (ROS) and expresses genes related to immune responses in zebrafish embryos [8]. Through docking studies with enzymes, Jayakanthan et al. [9] observed that anti-oxidant enzymes such as catalase (CAT) and superoxide dismutase (SOD) were strongly inhibited by 4-NP in the embryo. In Arabidopsis thaliana, a weed plant, 4-NP stimulates lipid peroxidation in leaves along with the induction of anti-oxidant enzymes such as ascorbate peroxidase, CAT, and SOD [10]. It has also been reported that 4-NP interferes with male reproductive function in mammals by producing ROS [11,12]. All these data obtained from different organisms collectively suggest that 4-NP might influence endogenous antioxidant systems in fish. Studies that have examined the toxicity of 4-NP in fish species are generally rare, and furthermore those on anti-oxidant systems are almost absent. Most of the ecological studies on 4-NP in fish have been limited to endocrine systems.
In view of the ubiquitous presence and ecotoxicological significance of 4-NP in aquatic environments, an assessment of the influence of this contaminant on the hepatic anti-oxidant systems of the Far Eastern catfish Silurus asotus was performed in this study. This specific catfish species is indigenous to Far Eastern Asian countries including Korea, China, Taiwan and Japan. As it is very well-known that 4-NP is widely present in most of the aquatic environments of Korea, the assessment of the adverse effects of this contaminant is highly meaningful. The significance of the observations reported in this study may be further highlighted by the fact that 4-NP levels used range within environmentally relevant low concentrations.
Materials and Methods
Experimental Fish and Chemicals
Far Eastern catfish were purchased from a local fish farm in Jeonbuk Province, Korea. Catfish were acclimatized under laboratory conditions maintained at 23±2°C under a 12:12 hours light/dark cycle. During acclimatization, fish were fed a basal diet (control diet in Table 1) twice daily at 2% body weight equally divided at 10:30 am and 4:30 pm. After acclimatization, fish of similar sizes (12 to 14 g) were selected for exposure exexperiments. The test aquaria (370-liter capacity rectangular tank composed of fortified reinforced fiber) were supplied with aerated and dechlorinated continuous flow-through water (flow rate=8 L/min). Sixty fish for each concentration level were used and 10 fish were sampled for biochemical analysis at weeks 0, 2, and 4 after exposure. For data not requiring fish euthanization (e.g., body weight measurement), all remaining fish at designated time points were used. Prior to euthanization, fish were anesthetized with neutralized MS-222 (Sigma-Aldrich, St. Louis, MO, USA). All procedures for fish handling were performed according to the Ethical Codes for Animal Experiments of Kunsan National University. Most of the other major chemicals were obtained from Sigma-Aldrich.

Effects of 4-nonylphenol (4-NP) administration on survival, body weight, hepatosomatic index (HSI) and feed intake in Far Eastern catfish
Diet Preparations
Experimental diets (Table 1) were formulated as described by Kim et al. [13] for this catfish species and 4-NP (CAS No. 104- 40-5, Sigma-Aldrich, >98%) was supplemented at 0.1 and 1.0 mg/kg diets. For this, 4-NP was first dissolved in absolute ethyl alcohol before mixing with a feed mixer. The mixture was extruded while moist through a 3-mm diameter extrusion module. The resulting pellets were freeze-dried to a moisture content of about 8.0%. Pellets were crumbled into particles, sieved to obtain appropriate sizes and stored at -70°C until fed. The control diet contained (in %): white fish meal 40, casein 13, beef tallow 5, corn oil 5, dextrin 27, α-cellulose 5.5, vitamin mix 2, mineral mix 2 and choline chloride 0.6 [13]. As the 4-NP content constituted a small portion of the diet, i.e., 0.01-0.1 mg/100 g diet in the 4-NP exposure groups, 4-NP was added to the control diet without further composition adjustments.
General Assessment of Toxic Signs
For the general assessment of toxic signs, mortality and body weight changes were measured during exposure at the indicated time points. Instantaneous maximum food intake was assessed by supplying the feed at two-fold the expected satiation level and unconsumed pellets was retrieved for weighing. The hepatosomatic index (HSI) was defined as the wet weight ratio between the hepatopancreas and whole body weight.
Preparation of Liver Tissues for Biochemical Analysis
At designated time points, livers were excised from anesthetized fish. The liver was then homogenized in four to five volumes of ice-cold tris buffer (50 mM, 1 mM EDTA, 1 mM dithiothreitol, 150 mM NaCl, pH 7.5) with several passes in a tissue homogenizer (X520; Ingenieurbüro CAT, Staufen, Germany). The homogenate was centrifuged (12000×g for 15 minutes, Hanil, Seoul, Korea) at 3°C and the supernatant was used for assays.
Lipid Peroxidation Levels
Levels of thiobarbituric acid-reactive substances (TBARS) were measured to estimate hepatic malondialdehyde (MDA) concentration by the method described by Ohkawa et al. [14]. An aliquot of liver homogenate (1.0 mL) was mixed with 2 mL of 0.1% thiobarbituric acid solution, and heated in a boiling water bath for 15 minutes. The reaction mixture was cooled down to room temperature and centrifuged at 1000×g for 10 minutes to obtain a pink colored supernatant. The absorbance of the supernatant was measured at 532 nm against reagent blanks. TBARS were quantified using a standard curve constructed with 1,1,3,3-tetraethoxypropane.
Reduced Glutathione Contents
The reduced glutathione (GSH) concentration was measured by a kinetic spectrophotometric method [15]. In short, 0.5 mL homogenate was precipitated with 5% trichloroacetic acid (TCA) to remove protein. After centrifugation, 100 μL of the clear supernatant was added to tubes containing 2 mL 5,5’-dithiobis(2-nitrobenzoic acid) (0.6 mM in 200 mM phosphate buffer, pH 8.0), and the mixture volume was brought to 4.0 mL with 200 mM phosphate buffer. Absorbance was read at 412 nm against a solution containing only TCA. Quantification was performed using standard GSH solutions.
Glutathione Reductase Activity
Glutathione reductase (GR) activity was measured at 340 nm with a slight modification of the method by Goldberg and Sparner [16]. The reaction mixture contained 50 mM imidazole, 0.3 mM Na2-EDTA, 0.1 mM NADPH and 2 mM oxidized GSH (GS-SG, pH 7.1).
Glutathione Peroxidase Activity
Glutathione peroxidase (GPx) activity was measured in accordance with Lawrence and Burk [17]. An aliquot of the hepatic supernatant (50 μL) was added to potassium phosphate buffer (50 mM) containing 1 mM EDTA and 1 mM Na3N. To the reaction tubes, 0.2 mM NADPH, 1 mM GSH, 1U GR, 0.25 mM H2O2 and 1.5 mM cumene peroxide (dissolved in 10% ethanol initially) were added and absorbance at 340 nm was kinetically recorded at 25°C.
Superoxide Dismutase Activity
SOD activity was measured according to Kakkar et al. [18]. The enzymatic reaction was initiated by incubating 1.0 mL of liver homogenate with nitroblue tetrazolium, NADH and phenazine methosulfate for 90 seconds at 30°C. The reaction was stopped by adding 100% glacial acetic acid. The color product produced was extracted with n-butanol and absorbance was measured at 520 nm.
Catalase Activity
CAT activity was assessed by the colorimetric method described by Sinha [19]. For this, 100 μL of tissue homogenate was allowed to degrade 200 mM H2O2 at a ratio of 1:4. The reaction was stopped at 30 seconds intervals by adding 2.0 mL dichromate- acetate solution (prepared at a 1:3 ratio). The reaction mixture was placed in a boiling water bath for 10 minutes followed by cooling. The developed colored substance was quantified at 620 nm using H2O2 standard solutions.
Protein Contents
The protein contents of the tissue assay samples were assessed according to the method of Bradford with bovine serum albumin as the standard [20].
Statistical Analysis
Data are expressed as mean±standard deviation. Significant differences among groups were determined using one-way analysis of variance followed by Duncan’s test for multiple comparisons [21]. Statistical significance was determined at p-value < 0.05.
Results
General Parameters
Table 1 shows the test group design and some parameters measured to estimate the signs of 4-NP induced general toxicity. There was one death out of 60 fish in the 0.1 mg/kg diet group; this occurred on day 24 of treatment. No mortality occurred in either control or 1.0 mg/kg diet group. There was a slight retardation of body weight increase after 4 weeks of 1.0 mg/kg administration.
Malondialdehyde Levels
The hepatic concentration of MDA, which reflects the degree of tissue lipid peroxidation, is illustrated in Figure 1A. There was no significant change in MDA in control catfish throughout the four-week exposure period. However, the MDA level was significantly elevated after two weeks by both 0.1 and 1.0 mg/kg 4-NP. These elevated levels were similarly maintained throughout four weeks.
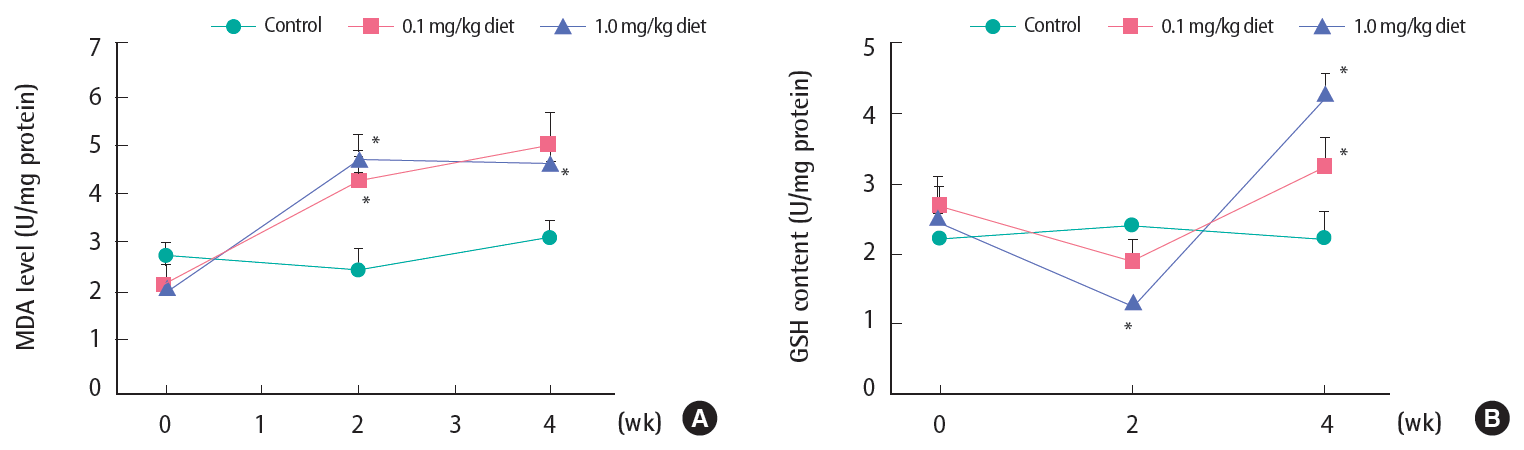
Changes in levels of hepatic (A) malondialdehyde (MDA) and the reduced form of (B) glutathione (GSH) after 4-nonylphenol exposure in Far Eastern catfish. Mean±standard deviation (n=10). Statistical comparison was made against control values at the corresponding time points. *p<0.05.
Figure 1B shows GSH levels during the four weeks of 4-NP exposure. In the control group, the levels were stable throughout the four-week experimental period. There was a clear tendency for a decrease after two weeks of exposure with the two 4-NP diets: however, statistical significance (p<0.05) was attained with 1.0 mg/kg but not with 0.1 mg/kg (p=0.07). At week 4, GSH levels increased even above the control levels with both the 0.1 and 1.0 mg/kg 4-NP diets.
Glutathione Reductase Activity
Figure 2 illustrates changes in GR activity, the enzyme that reduces the oxidized form of GSH (GS-SG) to the reduced one (GSH), thereby restoring the reduction potential of the cell. The observed pattern shows that there was no substantial change in control livers during the four-week study period. Upon exposure to 0.1 mg/kg 4-NP, there was no change in the activity within two weeks. However, it increased significantly after four weeks. With the higher level of 4-NP, 1.0 mg/kg, GR activity tended to increase from week 2 (statistically non-significant, p=0.064) and the level further increased after week 4.
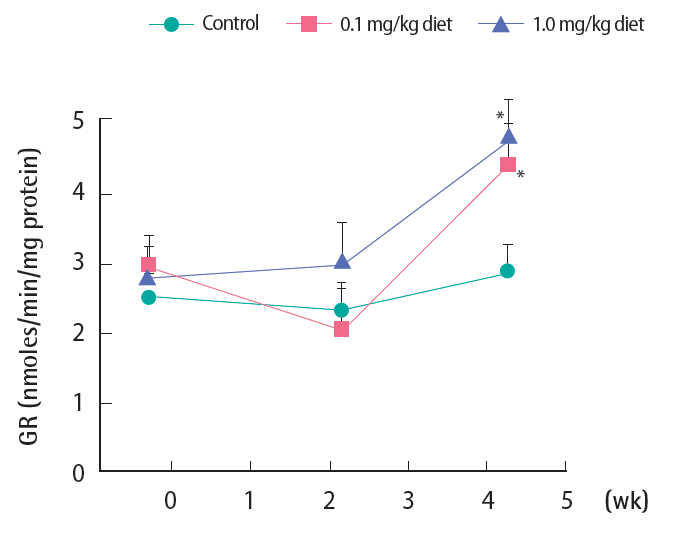
Activity of hepatic glutathione reductase (GR) following 4-nonylphenol exposure in Far Eastern catfish. Mean±standard deviation (n=10). Statistical significance was checked against control values at the corresponding time points. *p<0.05.
Glutathione Peroxidase Activity
GPx was significantly reduced with 4-NP treatment after two weeks at the two tested levels (Figure 3). Such significant decreases recovered completely to control levels by week 4.
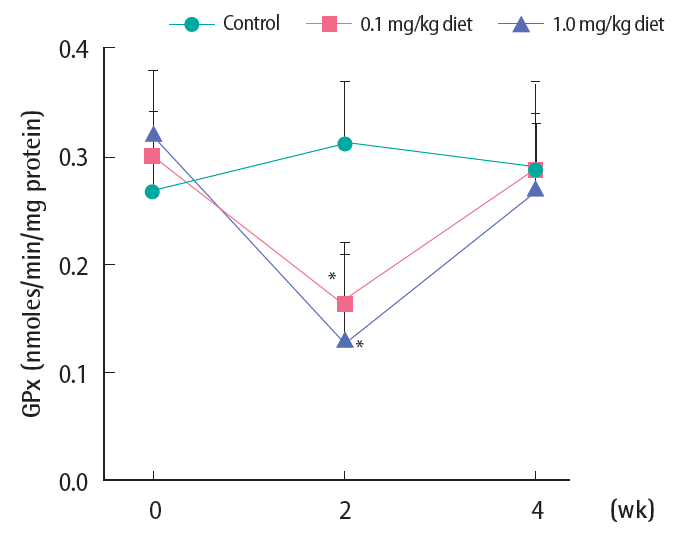
Activity of hepatic glutathione peroxidase (GPx) following 4-nonylphenol exposure in Far Eastern catfish. Mean±standard deviation (n=10). Statistical significance was checked against control values at the corresponding time points. *p<0.05.
Superoxide Dismutase Activity
Changes in SOD activity are illustrated in Figure 4. There was no significant change in SOD activity either in the control or 0.1 mg/kg diet fish groups. However with 1.0 mg/kg 4-NP, SOD increased significantly from week 2 and the elevation was maintained until week 4.
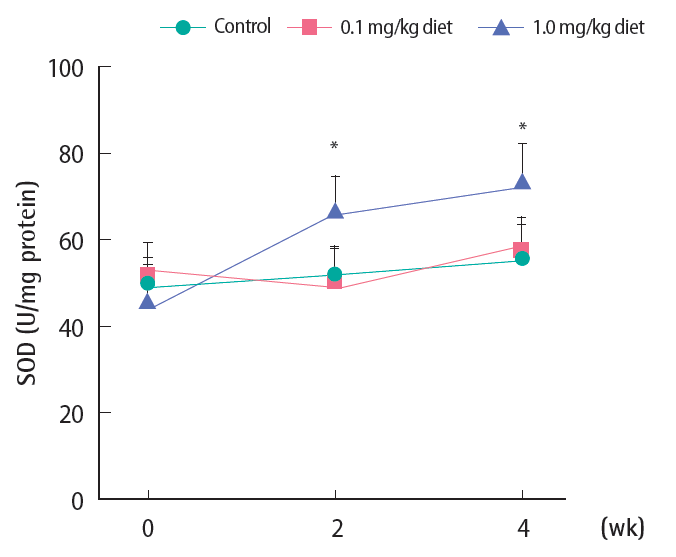
Activity of hepatic superoxide dismutase (SOD) following 4-nonylphenol exposure in Far Eastern catfish. Mean±standard deviation (n=10). Statistical significance was checked against control values at the corresponding time points. *p<0.05.
Catalase Activity
CAT activity did not change either in the control or 0.1 mg/kg diet groups throughout the four-week study period. At 1.0 mg/ kg 4-NP, however, a significant increase was noted after four weeks (Figure 5).
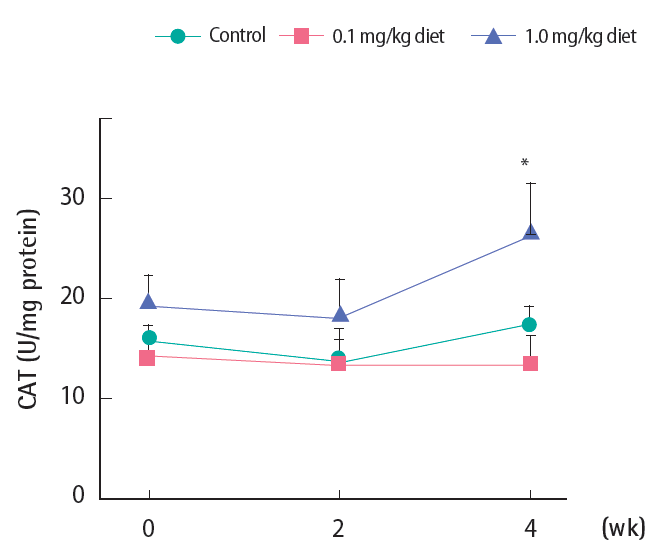
Activity of hepatic catalase (CAT) following 4-nonylphenol exposure to Far Eastern catfish. Mean±standard deviation (n=10). Statistical significance was checked against control values at the corresponding time points. *p<0.05.
Discussion
This study describes the pro-oxidant effects of 4-NP and the compensatory responses of hepatic anti-oxidant systems, possibly activated to alleviate oxidative stress in the Far Eastern catfish Silurus asotus.
With four weeks of exposure to 4-NP at 1.0 mg/kg, it was observed that normal body weight gain of the fish was slightly retarded. Body weight change is a rough but good indicator of general toxicity in an animal. From the data on instantaneous feed intake with 1.0 mg/kg 4-NP, which was not different from control, it seems that the reduced body weight gain was the result of reduced feed consumption. The HSI is used to assess hepatic tissue damage or the depletion of energy reserves in the liver. As HSI was not changed by 4-NP, it can be deduced that massive hepatic damage did not occur, even with the higher exposure level. These background data imply that influences identified in this study were quite specifically ascribable to anti-oxidant systems, rather than to the indirect consequences of the general toxicity of 4-NP.
Of all reported toxicities of 4-NP in animals, malfunction of the endocrine system is the best described [22]. As a possible mechanism for the adverse effects of 4-NP on male reproductive function, accelerated production of ROS was proposed in the mammalian epididymis [11]. In a study by Aly et al. [11], enzyme activities of SOD, CAT and GPx were all decreased in contrast to the increases in most enzymes observed in the current study. It is not clear yet what caused such a discrepancy. Reasons that can be considered include sensitivity differences between species or test dose differences. Very high doses (>100 mg/kg/d vs. 2-20 μg/kg/d) of 4-NP were used in the Aly’s study [11].
Overall observations from this study indicate that 4-NP is in fact a strong pro-oxidant, and that hepatic enzymes associated with GSH or other anti-oxidant systems seem to operate compensatory mechanisms to overcome the pro-oxidant burden. With the two weeks of exposure, there was a significance increase in lipid peroxidation products (MDA), equally at 4-NP levels of 0.1 and 1.0 mg/kg diet. Such accelerated lipid peroxides probably led to 4-NP level-dependent consumption of reduced GSH. After four weeks, GSH levels increased over the pre-exposure levels in both exposure groups, indicating the possible activation of compensatory GSH over-production. Interestingly, however, MDA levels at week 4 after 4-NP treatment at both levels did not return to their control levels. This may imply that GSH is not the sole anti-oxidant mechanism mobilized to scavenge 4-NP-induced lipid peroxides. Various anti-oxidant mechanisms, for example vitamin C, vitamin E, and α-lipoic acid have been reported to control pro-oxidants in cells [23].
It was observed that GR, the enzyme that regenerates GSH by reducing oxidized GSH (GS-SG), was increased within two weeks of exposure, at which time lipid peroxide levels were not recovered to the control state. The enzyme activity of GPx, an enzyme responsible for quenching lipid peroxides, was similarly stimulated by GR in the temporal sense. Other anti-oxidant enzymes examined in this study, SOD and CAT, were also elevated by 4-NP. However, these enzymes seem to be less sensitive to 4-NP than GSH-involving enzymes because those enzymes were stimulated at the 1.0 mg/kg level, but not at 0.1 mg/kg. Although GR helps produce GSH in principle, elevated enzyme activities of the enzyme will not always result in a net increase of GSH levels. Depending on the balance between the production and consumption of GSH, there could be a decrease in GSH levels even while the enzyme activity is higher [24]. This study did not examine the mechanism of activation of anti-oxidant enzymes after 4-NP administration in catfish. It is presumable, however, that 4-NP might have stimulated the expression of these enzymes. For example, Hernandez et al. [25] demonstrated that 4-NP induces the expression of a series of hepatic cytochrome P450s in mice after seven days of repeated administration. P450 aromatase, responsible for the biosynthesis of estrogens, was also found to be expressed in the whole body of juvenile zebrafish after three days of exposure [26]. In the same line, P450 enzyme genes were expressed when other fish were exposed to 4-NP [27,28].
The findings obtained in this study carry high significance because the 4-NP concentrations administered to our experimental fish were very low in comparison with those reported by others. For example, most studies have delivered 4-NP in the range of milligrams to several hundred milligrams per kilogram body weight [29]. In comparison, the current study tested 0.1 and 1.0 mg/kg diet which corresponds to 2 and 20 μg/kg body weight (Table 1). It is highly plausible that such low level exposure can occur through commercial fish feed or ambient culture water. It should be borne in mind that 4-NP is detected ubiquitously in wild fishes which could be major resources for commercial fish feeds [29,30].
MDA levels were elevated after two weeks, along with compensatory anti-oxidant enzyme systems. Such systems seem indicate the activation of anti-oxidative mechanisms over time, because MDA concentrations leveled off after four weeks. These biochemical alterations observed in Far Eastern catfish could be useful as biomarkers to deduce prior exposure to 4-NP. There was a clear difference in the kinetics and also the sensitivity for the activation of individual enzymes, with some parameters responding earlier than others.
Acknowledgements
This work was financially supported by the Faculty Research Funds of Kunsan National University in the fiscal year 2014.
Notes
The author has no conflict of interest with the material presented in this paper.